Biological mirrors
Biological mirrors are prevalent in biology. They are light-reflecting structures in organisms made of materials with high refractive index embedded in a low refractive index media like water or cytoplasm in cells. This structures -which can be extremely complex- are responsible for many of the structural colors that we find in nature.
The majority of frog structural colors are produced by skin cells that contain guanine crystals, known as iridophores. However, beyond skin cells, hundreds of treefrog species have bright white tissues covering their internal organs or muscles that broadly reflect both visible and NIR light.
The majority of frog structural colors are produced by skin cells that contain guanine crystals, known as iridophores. However, beyond skin cells, hundreds of treefrog species have bright white tissues covering their internal organs or muscles that broadly reflect both visible and NIR light.
We found that BBSs are responsible for the modulation of leaf-like green colors in treefrogs. These proteins create green by absorbing blue and red light. However, the colors we see are not only the result of light absorption; when pigments are overly concentrated, they actually remove additional light, making the animal darker. In order to control the brightness and saturation of the frogs, treefrogs need to be able to enhance the BBS's non-absorbed green section of the spectrum. This is where the underlying white mirrors in their organs and peritonea come into play. Biological mirrors are key in the creation of colors in biology, but our knowledge on the anatomy, biochemistry and optics of these internal mirrors is limited.
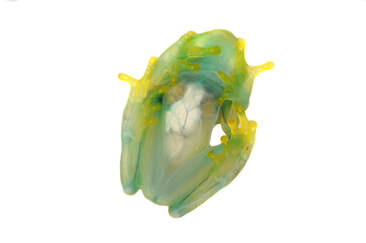
Our preliminary results showed that white peritonea contain millions of nanocrytals made of guanine/hypoxanthine, nitrogen-rich purines derived from DNA metabolism.

We are interested in the biochemical mechanisms of nitrogen metabolism that allow the deposition of crystals. We are also interested in how different structural organizations of these crystals create local variations in the backscatter properties of the tissues and ultimately at the whole organismal level. For this aim we use a combination of techniques from molecular biology and biochemistry, optics, microscopy, chemistry and computational approaches such as Monte Carlo.
Near-Infrared Reflectance: Convergent mechanisms of spectral red edge in frogs
Hundreds of treefrogs have a characteristic red edge in their reflectance spectra. Red edge is a steep increase in the amount of light that is reflected in the transition from the red to the infrared portion of the spectrum, and is a characteristic feature of foliage light reflectance. We demonstrated that the genesis of the red edge in numerous treefrogs depends on a variety of tissular, biochemical, and optical features, including translucent skins, high BBS concentrations, and biological mirrors. However, there are numerous additional species whose reflectance spectra converged to a leaf-like appearance, including the distinctive red edge, despite lacking some -or all- of these features. The Phyllomedusine frogs –such as the red-eye treefrogs or the monkey frogs, among many others– provide as an excellent example.
In the lab, we are interested in the convergent mechanisms that permit this trait. In collaboration with the Photoacoustic Imaging Lab at Duke University, we are using hyperspectral photoacoustic imaging to describe the absorption characteristics of pigmentary cells in the skin of near-infrared reflecting frogs in real life. Many pigments markedly change their optical properties in different chemical environments, for example, when the pigments are extracted, purified and solubilized in different solvents. The use of non-invasive imaging techniques allows us to investigate the optical properties of frogs' pigments in vivo which gives us a more realistic description of light absorption by the tissues. We use these results in combination with other optical measurements and various microscopies to account for scattering and incorporate these results in optical models to better understand the mechanisms that create the high NIR-reflecting frog phenoptypes.